|
|
Journal of Advanced Veterinary Research Volume 9, Issue 4, 2019, Pages: 151-160 www.advetresearch.com |
|


|
Inhibitory Activity of Silver Nanoparticles and Sodium Hypochlorite against Biofilm Produced by Salmonellae Isolated from Poultry Farms |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abd El-Moez A. Ismail, Saber A.H. Kotb, Israa M.A. Mohamed, Hosnia S. Abdel-Mohsein* |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Animal, Poultry Hygiene and Environ. Sanitation Dept. Faculty of Veterinary Medicine, Assiut University, Egypt. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Received: 7 July 019; Accepted: 22 September 2019 (*Corresponding author: hosnia18@aun.edu.eg) |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abstract |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Salmonella Typhimurium and Salmonella Enteritidis are among the predominant Salmonella serotypes in the Egyptian poultry farms. Salmonella has the ability to build up biofilms on a variety of surfaces. The antibiofilm activities of silver nanoparticles (AgNPs) and sodium hypochlorite (NaOCL) on prevention and controlling of biofilm by Salmonella spp. was estimated. Silver nanoparticles exhibited bactericidal activity against both S. Typhimurium and S. Enteritidis with MIC value at 15µg/ml, while, that of NaOCL was 1600 µg/ml. AgNPs (25 ug/ml) could inhibit biofilm formation at percentages of 84.96% and 78.85% against S. Typhimurium and S. Enteritidis, respectively. A percentage of 87 % biofilm removal by AgNPS after 3 h contact with the built-up biofilm produced by S. Typhimurium and S. Enteritidis was recorded. NaOCL (2200 µg/ml) exhibited inhibition percentages of biofilm formation at 83.89% and 75.76% against S. Typhimurium and S. Enteritidis, respectively. While, biofilm removal percentages after 2 h contact between NaOCL (2200 µg/ml) with the formed biofilm by S. Typhimurium and S. Enteritidis were 87.42% and 89.37%, respectively. It can be concluded that AgNPs and NaOCl were able to promote a significant reduction of biofilm formation by S. Typhimurium and S. Enteritidis. Also, AgNPs and NaOCl effectively oppress the mature biofilms formed and the antibiofilm efficiency increased with the increase of contact time with the biofilms. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Keywords: Biofilm; Poultry farms; Salmonella; Silver nanoparticles; Sodium hypochlorite |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Introduction |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bacterial biofilm is an association of micro-organisms within a self-produced matrix of extracellular polymeric substance, in which bacterial cells adhere to each other on living or non-living surfaces. It is infectious in nature and can result in various infections. Moreover, biofilm formation protects bacterial cells against a wide range of challenges including UV light radiation, pH and osmotic changes, dehydration, host immune responses, antimicrobial agents and disinfectants (Nilsson et al., 2011; Bogino et al., 2013). The established biofilms can tolerate antimicrobial agents at concentrations of 10-1000 times that needed to kill planktonic bacteria and also have an extraordinary resistance to phagocytosis, which make it difficult to eradicate biofilms from living hosts (Cos et al., 2010). Therefore, a greater understanding of bacterial biofilm is required for the development of novel, effective control strategies that can overcome the drawbacks of the current control ones. One of these novel alternatives could be metal-based nanoparticles that have been utilized in several applications due to their fast and broad antibacterial activity as well as low production costs (Fabrega et al., 2011). The antimicrobial activity of silver nanoparticles in reducing the growth of different microorganisms is well documented (Li et al., 2006; Kanematsu et al., 2009). Silver nanoparticles (AgNPs) are now considered one of the most promising strategies to control bacterial infections. The bactericidal efficiency of AgNPs has been attributed to their small size and high surface to volume ratio, which allows them to interact closely with microbial membranes (Morones et al., 2005; Rai et al., 2009). Furthermore, AgNPs have not been shown to cause bacterial resistance because silver nanoparticles exert their antibacterial effects at several sites (Shrivastava et al., 2007). Smaller AgNPs can reduce more biomass and viability of biofilms, due to better penetration into the exopolysaccharide (EPS) matrix (Habash et al., 2014). Moreover, the anti-biofilm activity of AgNPs could be due to inhibition of EPS synthesis in bacteria, which limits the biofilm formation (McLaughlin-Borlace et al., 1998). Sodium hypochlorite (NaOCl) is one of the most effective disinfectants against biofilms (Pui et al., 2011; Rodrigues et al., 2011); with the ability to eradicate biofilms at concentrations as low as 3.125 mg per ml (Rodrigues et al., 2011). During treatment, sodium hypochlorite decomposes to sodium hydroxide and hypochlorite, which is a strong oxidizing agent (Tote et al., 2010) that act by oxidizing the bacterial cell membrane resulting in cell lysis and death (Maris, 1995; Hawkins and Davies, 1999). In Egypt, poultry production represents one of the most important economic and animal protein sources for human, however this sector faces many problems and challenges. One of these problems is the infection with various microbes such as Salmonella. Salmonella enterica serovar Typhimurium and Salmonella enterica serovar Enteritidis have been identified as the predominant serotypes present in Egyptian poultry farms (Abd El-Ghany et al., 2012). There are many reports regarding the development of biofilms by Salmonella on a variety of biotic and abiotic surfaces, where they allow Salmonella to survive and spread in the environment (Janssens et al., 2008). Furthermore, Salmonella contamination on farms may come from a variety of sources, including wild animals, rodents, insects, feed and human (Meerburg and Kijlstra, 2007; Carrique-Mas et al., 2009; Hilbert et al., 2012). Control of Salmonella is a challenge to public health because of their emergence/ re-emergence and high mutation rate, with antibiotic resistance in both developed and developing countries (Jassim and Limoges, 2017). Salmonella found in biofilms showed a higher tolerance to antibiotics and was less susceptible to disinfectants than planktonic Salmonella (Pui et al., 2011; Sheffield and Crippen, 2012). The use of glutaraldehyde, formaldehyde and peroxygen at a concentration of 1% in field conditions is insufficient to eradicate Salmonella biofilms (Marin et al., 2009). High prevalence of Salmonella spp. in water troughs swabs and water was reported by Ogden et al. (2007) and Marin et al. (2009), where many strains of Salmonellae were able to produce biofilm in water tanks, drinker lines and stainless-steel surfaces, and continuously release planktonic cells in water. Therefore, this biofilm is one of the potential and continuous sources of infection because it helps salmonellosis to be transferred to the entire flock. So, to achieve efficient control of Salmonella infection in poultry farms there is a need for an effective method for removal and prevention of biofilm formation on these surfaces. The misuse of antibiotics and different antimicrobials in the Egyptian poultry industry led to the emergence of resistant strains that are difficult to eradicate. Therefore, we need to evaluate the activity of other alternative agents against the strains isolated from Egyptian poultry farms to be able to achieve good hygienic measures and control biofilm producing bacteria in both poultry farms as well as human establishment. The objective of the current study was to evaluate the effectiveness of AgNPs and NaOCL on prevention and controlling of biofilm formation by Salmonella spp. isolated from Egyptian poultry farms. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Materials and methods |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bacterial strains The bacterial strains used in the current study were isolated from 11 poultry farms located in Assiut Governorate, Egypt. A total of 600 samples were collected from the poultry farms including litter, feed, water samples, as well as water troughs, cloacal and wall swabs. Salmonella detection was carried out according to standard methods (Anon, 2007), the samples were initially transferred into buffered peptone as a pre-enrichment medium and incubated for 18-20 h at 37°C then 100 µl of the pre-enriched cultures was transferred to Rappaport-Vassiliadis broth (Biolife EM1804) and incubated at 42°C. After 24 h of incubation, one loopful of enriched broths was streaked onto plates of xylose lysine deoxycholate agar (Biolife EG8702) and incubated at 37°C for 24-48 h. The plates were examined for the presence of typical colonies of Salmonella spp. i.e. red colonies with black centers (Antunes et al., 2003). Pure colonies were subjected to identification using gram staining, biochemical reactions and finally multiplex PCR for detection of S. Typhimurium and S. Enteritidis that was performed in Molecular Biology Unit, Assiut University, Egypt. S. Typhimurium strain was obtained from Animal Health Research Institute, Giza, Egypt and served as a positive control during multiplex PCR. As shown in Table 1, the primers’ sets used were ST11 and ST15 as a universal gene for Salmonella spp., SI and S4 for S. Enteritidis, and Fli15 and Typ04 for S. Typhimurium according to Soumet et al. (1999). Extraction was carried out using Patho Gene DNA/RNA extraction kit (iNtRON Biotechnology) according to the manufacture’ instructions. Multiplex PCR reaction was done in a total volume of 22 µl containing 11µl PCR master mix, 1µl of each primer and 5 µl of the extracted DNA. The thermocycler conditions included initial denaturation at 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 48°C for 1 min and 72°C for 1 min with final extension at 72°C for 10 min. After amplification, 1% agarose was used for electrophoresis. Table 1. Oligonucleotide sequences used to detect Salmonella spp. (Soumet et al., 1999)
The ability of each bacterial strain to produce biofilm in pure culture was determined using the tissue culture plate method according to Coffey and Anderson (2014). One to two colonies from fresh overnight cultured agar plates of each strain were transferred to 5 ml tryptic soy broth (TSB) and the bacteria was allowed to grow at 37 oC for 18 h. Then a 1:100 dilution was prepared in TSB using 0% glucose and 2% glucose as a supplement (Ganjali Dashti et al., 2016) and following thorough mixing, 100ul was transferred to each well in 96 microtiter tissue culture plates then incubated at 28oC without shaking for 48 h. After that the liquid media from each well was gently removed and the wells were washed three times with distilled water to remove the free-living bacterial cells that do not contribute to biofilm formation. The wells were stained with 125 µl of 0.1% crystal violet for 15- 30 min to confirm the presence of biofilm. Excess stain was rinsed off by thorough washing with distilled water three times, and then the plates were kept for 30 min till dryness. The microtiter plate was examined for the presence of purple ring that indicating the biofilm formation. For biofilm quantification 150 µl 30% acetic acids were added to each well and kept for 10 minutes and the contents of each well were mixed by repeated pipetting, and then 125 μl of the solubilized solution was transferred to a new well of a new optically clear flat bottom 96-well plate. The optical density (OD) of each well was measured at a wavelength 545 nm using Microtiter plate reader (Model Start Fax- 2100, Awareness Technology INC). Efficiency of AgNPs and NaOCL against S. Enteritidis and S. Typhimurium biofilm Preparation of silver nanoparticles (AgNPs) Stable AgNPs less than 100 nm was synthesized in a typical one-step protocol according to Vigneshwaran et al. (2006); where, one gram of soluble starch was added to 100 ml of deionized water and heated till complete dissolution, then 1 ml of a 100 mM aqueous solution of silver nitrate (AgNO3), was added and stirred well. This mixture was put into dark glass bottle and kept in an autoclave for 5 min at 121oC. The resulting solution was clear yellow in color indicating the formation of silver nanoparticles. After preparation of silver nanoparticles, the stock solution was kept away from direct sunlight at room temperature. The concentration and size of the particles were measured before its use. The total concentration of AgNPs was measured by Graphite Furnace Atomic Absorption (Model 210VGP) at the Faculty of Science, Assiut University, Egypt. The size of the particles was measured by Transmission Electron Microscope (TEM) (JEOL-JEM- 100CX II) at Electron Microscopy Unit, Assiut University, Egypt. Determination of minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of AgNPs and NaOCL The MIC of AgNPs and Naocl against S. Typhimurium and S. Enteritidis was measured by standard broth macro-dilution method as described by Sharma et al. (2015). Briefly, 1-2 overnight fresh colonies were added to 7 ml TSB and incubated for 18 h at 37oC. Different concentrations of AgNPs (5, 10, 15, 20, 25, 30 μg/ml) and NaOCL (5% available chlorine) (1000, 1200, 1400, 1600, 1800, 2000, 2200, 2400, 2600 and 2800 μg/ml) were separately added to each tube containing 5 ml TSB and finally an aliquot of bacterial culture (5×105 CFU ml-1) were added separately to these tubes. After that, the tubes were incubated at 37˚C for 18 h. In the control positive set, only the organisms were grown in absence of AgNPs and NaOCL, while tubes that contain TSB plus AgNPs and NaOCL in absence of inoculum were considered the control negative set. The MIC was considered as the lowest concentration of AgNPs and NaOCL in which there would be no visible bacterial growth after 18 h of incubation at 37˚C. To determine the MBC, the bacterial broth at the lowest concentration, which give no visible turbidity as well as at two higher concentrations were sub-cultured to sterile petri dishes containing standard plates count agar in duplicated to assure the absence of colony forming units (CFU). The petri dishes without any bacterial colonies were considered positive for the MBC. Prevention and eradication of Salmonella biofilm The ability of AgNPs and NaOCl to prevent biofilm formation and eradicate the established biofilm produced by Salmonellae was estimated according to Abidi et al. (2014). A total number of 12 strains including S. Typhimurium (n.=10) and S. Enteritidis (n.=2) were used in this experiment. Glucose supplement (2%) was applied during bacterial growth for biofilm production. At first the anti-biofilm activity of AgNPs and NaOCl was evaluated during bacterial incubation; while the biofilm was being created. Briefly, 1-2 colonies from fresh overnight cultured agar plates of each strain were transferred to 5 ml TSB and allowed to grow at 37oC for 18 h. Then the culture was diluted 1:100 in TSB with glucose supplement and 100 μl of diluted culture was pipetted in microtiter plate wells. After that, 100μl of each concentration of AgNPs or NaOCl was added into three wells and then plate was covered and incubated at 28oC for 48 h. Each bacterial strain was treated with four concentrations of AgNPs (10, 15, 20, and 25μg/ml) and four concentrations of NaOCL (1600, 1800, 2000 and 2200 μg/ml). Furthermore, for each strain, three wells of the 96-well flat bottom microtiter plate were inoculated with bacterial inoculums without treatments (positive control) and another three wells for negative control (treatment with TSB only). Concerning the second experiment, anti-biofilm activity of AgNPs and NaOCl was estimated after incubation; on the established biofilm. Briefly, microtiter plates were inoculated as mentioned above and incubated for biofilm production. After incubation, plates were washed with sterile water to remove planktonic cells after that 200 μl of each tested agent (25 μg/ml AgNPs and 2200 μg/ml NaOCL) was transferred into each well, with exception of blank and positive control wells. The plates were incubated for different contact times, namely (30, 60, 90, 120, 150 and 180 min) for AgNPs and (15, 30, 45, 60, 90 and 120 min) for NaOCL. At the end of each contact time, AgNPs and NaOCl were quenched by adding 5 g/l sodium thiosulfate (Na2S2O3) to stop their antimicrobial reaction as described in the European Quality Standards (NEN-EN 1276, 1997). Following the desired incubation time, the planktonic bacterial cells were removed from the microtiter plates and the wells were washed several times. Subsequently, for biofilm staining, 125 ul of 0.1% crystal violet solution was added to each well and incubated for 10 min at room temperature. Following incubation, the stain was removed, and plates were washed with vigorous shaking to remove all liquid. Subsequently, the plates were inverted and vigorously tapped on paper towels to remove all the contents and left to air dry. Finally, the dye was solubilized by adding 150 μl of 30% acetic acid to each well of the plate, and the plate was incubated for 10-15 minutes at room temperature. In the next step, contents of each well were mixed by repeated pipetting and then 125 μl of the crystal violet-acetic acid solution was transferred from each well to a separate well of a new optically clear flat- bottom 96-well plate. Optical density (OD) of each of these 125-μl samples was measured at a wavelength 545 nm., and to measure the antibiofilm efficacy; the reduction/ removal percentages were calculated using the following equation according to Abidi et al. (2014): Reduction/Removal Percentage = [(C-B) - (T-B) / (C-B)] *100% Where B = absorbance of blank (no biofilm, no treatment). C = absorbance of control (biofilm, no treatment). T = absorbance of test (biofilm and treatment). Statistical Analysis Statistical analysis of data was carried by using SPSS software version 17. The data was subjected to analyses of variance using the ANOVA procedure and General Linear Models Procedure (GLM procedure) of SPSS software. The results of optical densities were presented as mean and standard deviations (SD) for each variable. Significant differences between mean values were tested using Duncan's multiple range test. P-value consider statistically significant when P < 0.05. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Results |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
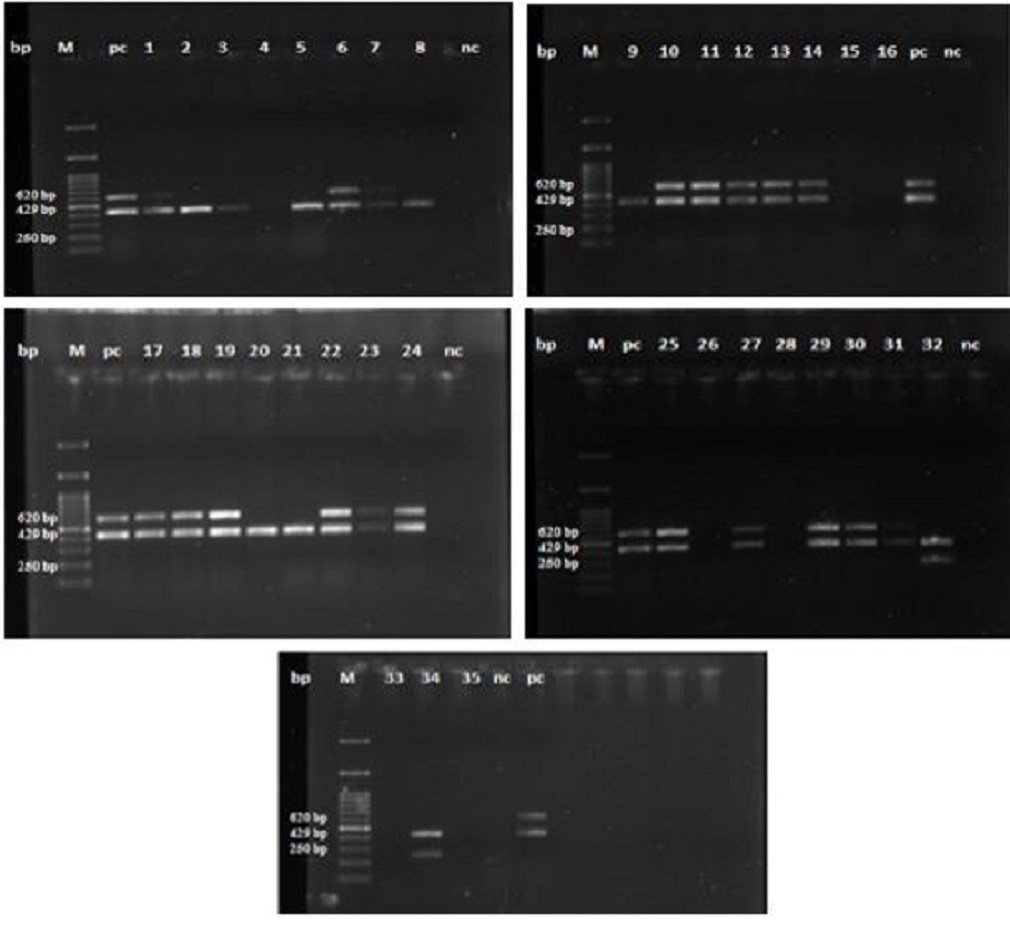
Isolation of Salmonella from poultry farms and the ability to build up biofilm Salmonellae were isolated from 11 broiler and layer farms in Assiut Governorate, Egypt. 35 biochemically positive Salmonella strains were subjected to molecular identification by multiplex PCR, from which 19 strains were positive for S. Typhimurium and two strains were positive for S. Enteritidis. Data illustrated in Fig. 1, represents the molecular identification of S. Typhimurium and S. Enteritidis by multiplex PCR. The data presented in Table 2, revealed that 54.29% of the strains were positive for S. Typhimurium with the highest percentage (26.32%) was recorded for litter samples followed by feed and water troughs swabs. Furthermore, 5.71% of the examined strains were positive for S. Enteritidis, representing 50 % for each cloacal and wall’ swab.
Fig. 1. Multiplex PCR identifying S. Typhimurium and S. Enteritidis. M:100 bp DNA marker; pc: positive control; nc: negative control; Lanes 2.3, 5, 8, 9, 20 and 21 are positive amplification of 429 bp fragments of Salmonella spp., Lanes 1, 6, 7, 10, 11, 12, 13, 14, 17, 18, 19, 22, 23, 24, 25, 27, 29, 30 and 31 are positive amplification of 620 bp fragments of Salmonella Typhimurium. Lanes 32 and 34 are positive amplification of 250 bp fragments of Salmonella Enteritidis. The ability of 21 Salmonella strains (19 S. Typhimurium and 2 S. Enteritidis strains) for biofilm formation was checked and the result revealed 10 strains S. Typhimurium and the 2 strains S. Enteritidis were able to produce biofilm. Table 2. Molecular identification of S. Typhimurium and S. Enteritidis by multiplex PCR
MIC and MBC of AgNPs or NaOCl on S. Typhimurium and S. Enteritidis As shown in Fig. 2, the size of AgNPs used in the current study was ranged from 7.55 to 18.9 nm with spherical shape. AgNPs exhibited bactericidal activity against S. Typhimurium and S. Enteritidis where the MIC value was 15µg/ml. While, the lowest concentration that prevent the bacterial growth on plate count agar (MBC) was 20 µg/ml. MIC and MBC of NaOCl on both S. Typhimurium and S. Enteritidis was 1600 µg/ml and 2000 µg/ml, respectively.
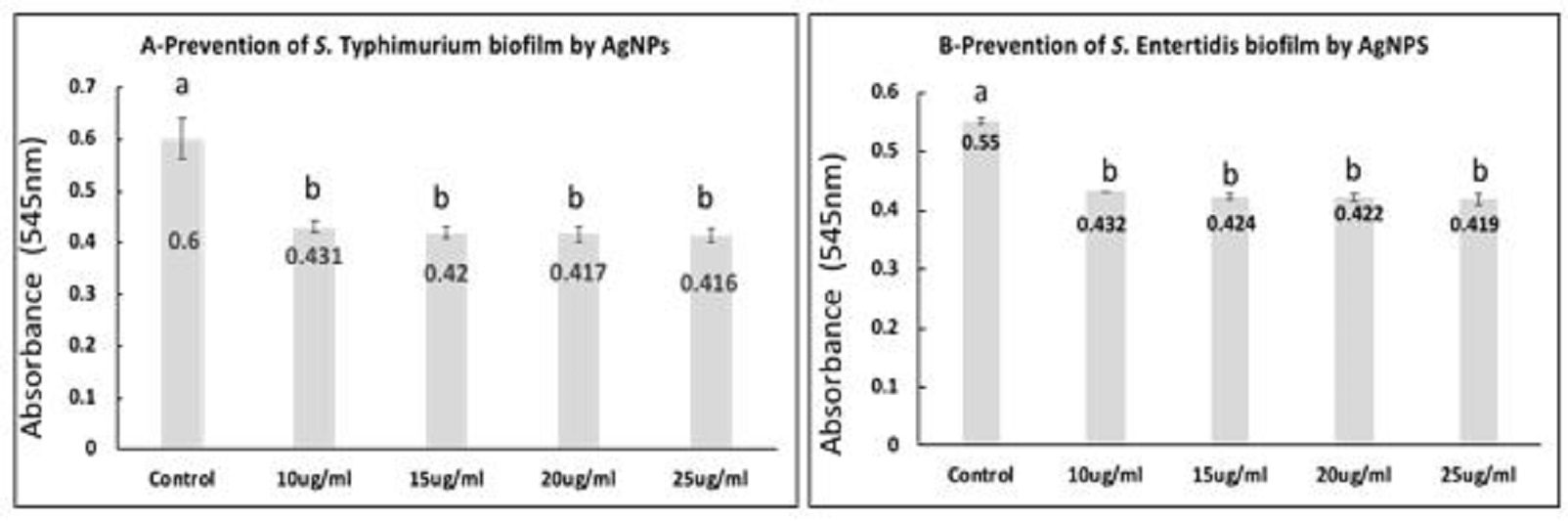
Fig. 2. Transmission Electron Microscope of silver nanoparticles (AgNPs) showing AgNPS spherical in shape with size ranged from 7.55 to 18.9 nm. Prevention of S. Typhimurium and S. Enteritidis biofilm formation in the presence of AgNPs or NaOCL As shown in Fig. 3A and B, the mean values of optical densities of bacterial cultures from quantified biofilm formed by S. Typhimurium and S. Enteritidis were significantly reduced in all AgNPs exposed bacterial cells at concentration of 10, 15, 20, 25 µg/ml when compared with the control (P<0.05). Furthermore, there were no significant differences between the mean values of optical densities of bacterial culture exposed to the various concentrations of silver nanoparticles. Moreover, the highest inhibition percentages; 84.96% and 78.85% of biofilm formation by S. Typhimurium and S. Enteritidis were observed when the bacterial cells treated with 25 µg/ml of AgNPs, respectively.
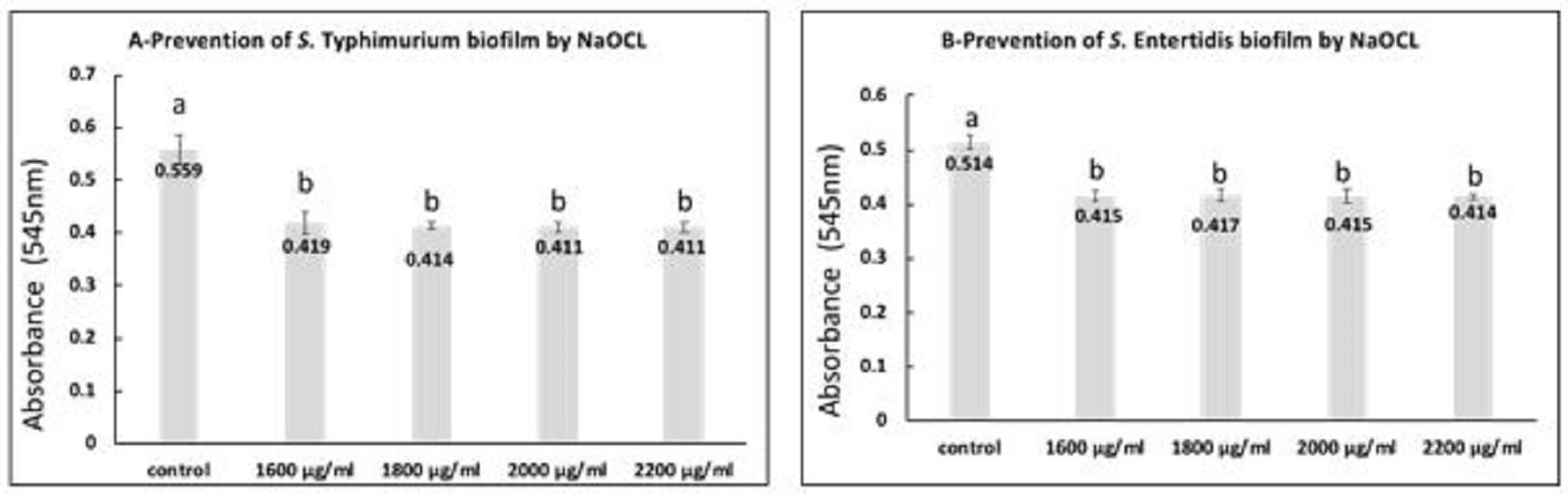
Fig. 3. S. Typhimurium (A) and S. Enteritidis (B) biofilms after 48 h incubation with the exposure to different concentrations of AgNPs. a,b: means with different letters are significantly different. The obtained data revealed that the mean values of optical densities of bacterial culture from the formed biofilm by S. Typhimurium and S. Enteritidis were significantly reduced in all NaOCL exposed bacterial culture when compared with the control (P<0.05) as presented in Fig. 4A, B. Furthermore, there were no significant differences among the mean values of optical densities of bacterial culture subjected to various concentration of NaOCL. When calculating the inhibition percentages of biofilm formation by S. Typhimurium and S. Enteritidis treated with 1600 µg/ml, 1800 µg/ml, 2000 µg/ml, 2200 µg/ml NaOCL, the highest percentages; 83.89% and 75.76% were observed at the highest used concentration, respectively.
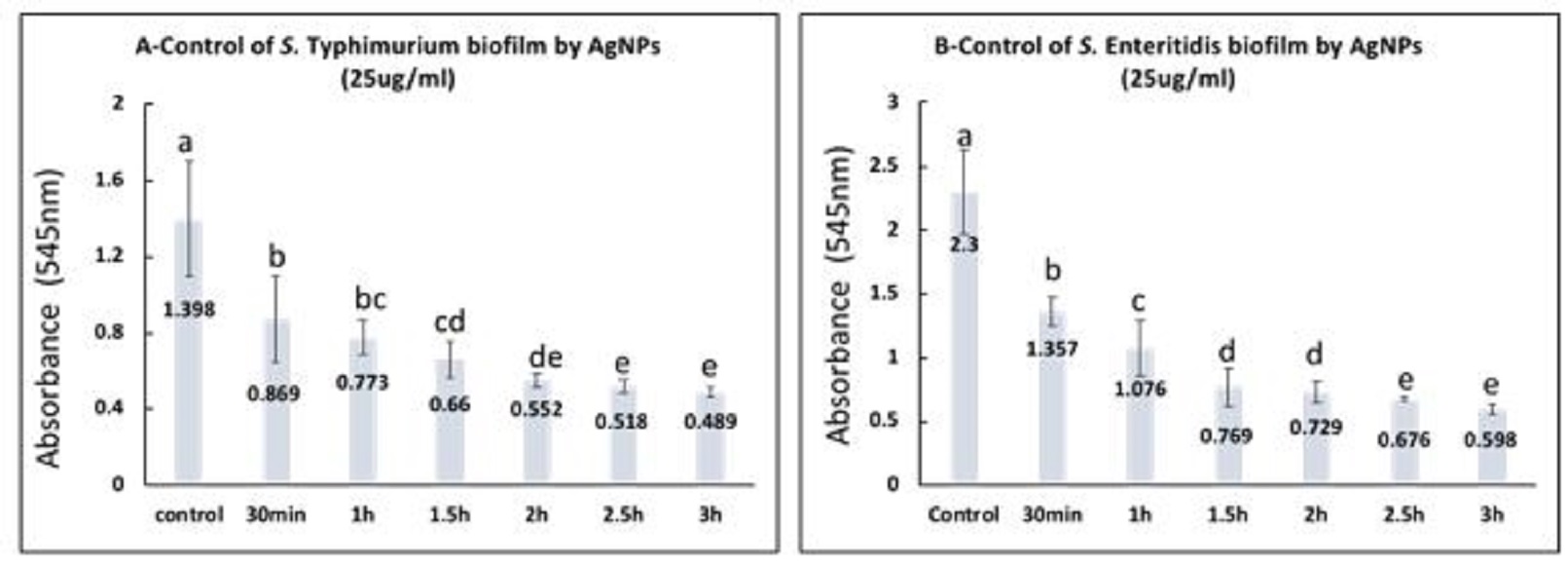
Fig. 4. S. Typhimurium (A) and S. Enteritidis (B) biofilms after 48 h incubation with the presence of different concentrations of NaOCL. a,b: means with different letters are significantly different. Control of biofilm formed by S. Typhimurium and S. Enteritidis by AgNPs (25 µg/ml) or NaOCL (2200 µg/ml) S. Typhimurium and S. Enteritidis established biofilms were exposed to AgNPs (25 µg/ml) for different contact times and the obtained data revealed that there were significant differences between the effect of AgNPs at all contact times and the control (P<0.05) as presented in Fig. 5. Moreover, there were significant differences between the mean values of optical densities for S. Typhimurium at 2.5 h, 3 h and the mean values at 30 min., 1 h, 1.5 h, and there were significant differences between the mean values at 2 h and the means at 30 min, and 1 h (P<0.05). The percentage of biofilm removal by 25 ug/ml AgNPS were 50.85%, 60.20%, 71%, 81.41%, 84.60% and 87.43% after contact time of 30 min, 1 h, 1.5 h, 2 h, 2.5 h, 3 h, respectively. For S. Enteritidis the results revealed that the biofilm removal percentage after the examined contact times were 48.70%, 63.15%, 78.93%, 80.94%, 83.63% and 87.67 %, respectively. Additionally, there were significant differences between all contact times and the control (P<0.05). Meanwhile, there were significant differences between the mean values at 2.5 h, 3 h, and the mean at 30 min, 1h, 1.5h, 2h, and there were significant differences between the mean at 1.5 h, 2 h, and that at 30 min (P<0.05).
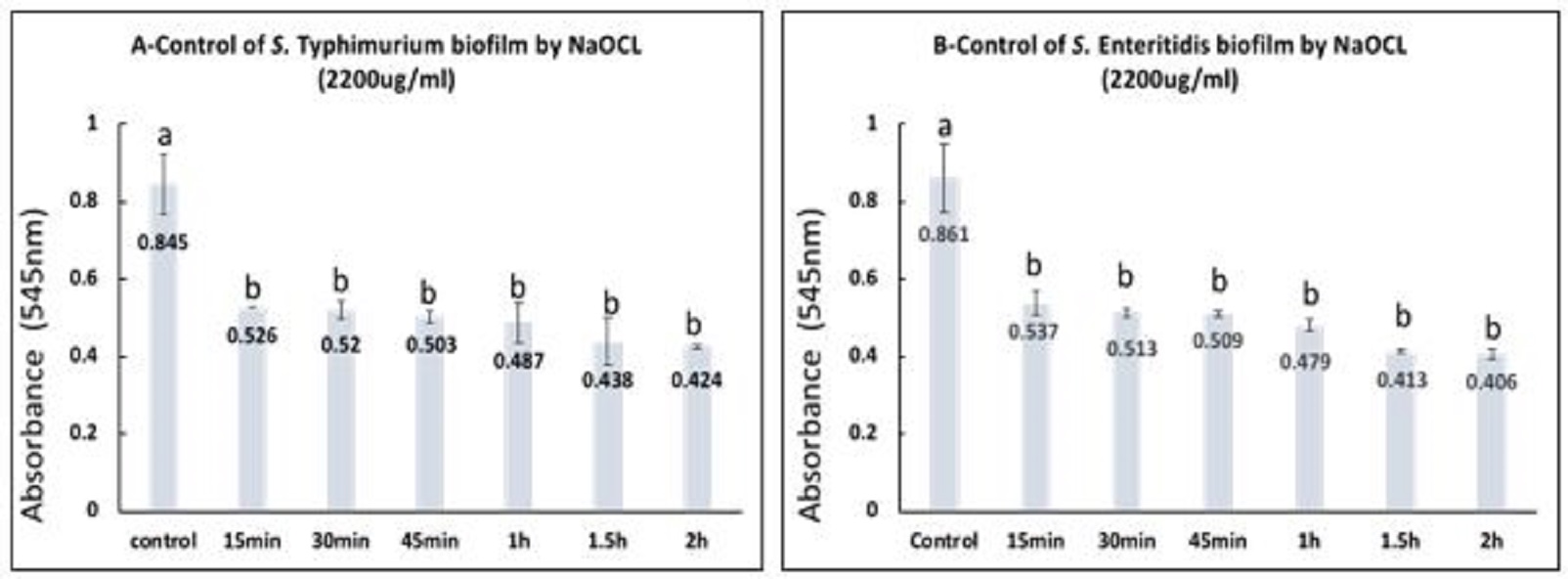
Fig. 5. Control of S. Typhimurium (A) and S. Enteritidis biofilms (B) by treatment with 25ug/mlAgNPs at different contact times. a,b,c,d,e: means with different letters significantly different. The percentages of biofilm removal by 2200µg/ml NaOCL after various contact times for S. Typhimurium and S. Enteritidis were estimated with the highest percentage 87.42% and 89.37% after 2 h contact, respectively. The statistical analysis of data for both S. Typhimurium and S. Enteritidis showed that there were significant differences between all contact times and the control (P<0.05), however, there were no significant differences among the different contact times as represented in Fig. 6 A, B. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fig. 6. Control of S. Typhimurium (A) and S. Enteritidis biofilms (B) by treatment with 2200ug/ml NaOCL at different contact times. a,b: means with different letters are significantly different. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Discussion |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Salmonella remains an important concern in food processing environments as it causes salmonellosis, a major public health problem throughout the world (Anonymous, 2004). S. Typhimurium and S. Enteritidis were the most frequently identified Salmonella serovars in the Egyptian poultry farms and their surrounding environment (Abd El-Ghany et al. 2012; Oliveira et al., 2012), moreover, they were the predominant serotypes contributing biofilm production (Marin et al., 2009; Cevallos et al., 2012), for these reasons and from the hygienic point of view, these serovars were selected to study the antibiofilm activity of silver nanoparticles and sodium hypochlorite. The obtained results revealed high prevalence of Salmonella spp. in litter samples, followed by water troughs swabs, cloacal swabs, feed samples, water samples and finally wall swabs. This finding agreed with Rodriguez et al. (2006), who reported that the prevalence of Salmonella in farm environments ranges from 10 to 26%. This can be explained by that faeces, soil, crevices, dusts, manure, litter, feeders and drinkers in farms and their environment harbor Salmonella, which increase the rate of contamination (Mallinson et al., 2000; Wales et al., 2006). Moreover, high prevalence of Salmonella spp. in litter is due to shedding of Salmonella from the intestinal tract of birds, which contaminates litter, bedding and feedstuff (Rodriguez et al., 2006; Hoelzer et al., 2011). The high prevalence of Salmonella spp. in water troughs swabs and water agreed with Murray (2000), who reported that Salmonella survives in water up to 56 days. Which may be attributed to failure to clean contaminated water or the water equipment after the removal of infected flock. Additionally, many strains of Salmonellae were able to produce biofilm in water tanks and drinker lines (Wilks et al., 2006; Ogden et al., 2007; Marin et al., 2009). Therefore, water is one of the potential sources of poultry infection because it helps salmonellosis to be transferred to the entire flock of the farm. The ability of 21 Salmonella strains (S. Typhimurium (n.=19) and S. Enteritidis (n.=2) strains) for biofilm formation was checked and the result revealed 10 strains S. Typhimurium and the two strains S. Enteritidis were able to produce biofilm. Addition of glucose during biofilm production is much more supportive in biofilm formation as glucose provided the basic carbon units and the nutrient broth supplied basic support for cell survival in biofilm (Coffey and Anderson, 2014; Ganjali Dashti et al., 2016). Salmonella is a hardy pathogen able to survive in many different environments (Carrique-Mas et al., 2009; Hilbert et al., 2012). In natural environments, bacteria exist in two forms; planktonic free bacteria are important for proliferation and spread of bacteria, or attached populations are necessary to allow the bacterial population to persist. During the last decades, it has become clear that bacteria, including food borne pathogens such as Salmonella enterica, grow predominantly as biofilms (Shemesh et al., 2007). Biofilm is a problem to the poultry industry, it is usually found in water systems of poultry farm and stainless steel in poultry processing plants and act as sources of cross-contamination (Wilks et al., 2006). Salmonellosis in chicken has been traced from biofilm associated with water tanks and drinker lines (Marin et al., 2009). Moreover, the transmission of S. typhimurium between hosts, their survival in the environment and persistence in the host are facilitated by the ability to form biofilms (Cevallos et al., 2012), furthermore, bacterial biofilms are more resistant to disinfectants (Wong et al., 2010). From all these, we need to change how we deal with micro-organisms on dry surfaces, biofilm (Yezli and Otter, 2012; Donskey, 2013). MIC and MBC value of AgNPs was 15 and 20 µg/ml, respectively. The obtained result comes agreed with Irayyif et al. (2015), who found that AgNPs at 10 nm size were effective towards Salmonella. The bactericidal effect of AgNPs depends on the size and shape of the particles, the smaller particle size increases the specific surface area of nanoparticles, allowing greater number of particles attached to a bacterial cell (Pal et al., 2007, Panáček et al. 2006). Loo et al. (2018) recorded that AgNPs; synthesized using pu-erh tea leaves extract with particle size of 4.06 nm, showed MIC against S. Typhimurium, and S. Enteritidis at 3.9 and MBC at 7.8, 3.9 μg/mL, respectively. MIC and MBC obtained from this study were higher than that recorded by Samberg et al. (2011), where MIC and MBC were 3-4 and 6-12 μg/ml AgNPs (20, 50, 80 um diameter) against Salmonella spp., respectively. Also, no visible growth was observed at concentration 6 μg/ml and MBC was 8 ug/ml AgNPs (75-86 nm diamter) (Gnanadhas et al. 2013). The size of AgNPs in this study is much smaller than that recorded by Samberg et al. (2011) and Gnanadhas et al. (2013). The antibacterial efficacy of AgNPs depends not only on the different particle sizes but also on surface conditions, furthermore, the surface condition of the particles is directly related to the synthesis method (Samberg et al., 2011). This may explain why result from this study differs from that of other authors that may be due to the different methods of synthesis. The antimicrobial action of AgNPs may be due to: (1) binding of Ag+1 that prevent the uptake of essential nutrients to bacterial cell, leading to cell death, (2) Ag+1 entry into the cell by competitive binding with essential heavy metals such as Ca2+, Mn2+ and Mg2+, or (3) their transport and irreversible accumulation in the cell could occur by complexation with substrates. Finally, Ag+1 could inhibit respiration, or bind and condense DNA once inside (Holt and Bard, 2005). In the end, the antimicrobial activity is likely caused by a synergistic effect between the binding of Ag ions to the cell wall, their uptake and subsequent accumulation in the cell, and their interference with critical biomolecules within the cell. MIC and MBC of NaOCl was 1600 µg/ml and 2000 µg/ml, respectively. These inhibitory levels were lower than that reported by Capita et al. (2017), who reported MIC at 6.0 mg/ml. The obtained levels were much higher than that recorded by Corcoran et al. (2014) and Espigares et al. (2006), who recorded inhibitory levels of 250 and 33-41µg/ml, respectively. The difference in the inhibitory levels may be attributed to difference in the available chlorine, contact time, method of analysis and different bacterial isolates. Furthermore, the physico-chemical characteristics of NaOCl as pH and surface tension are important for its mechanism of action and antimicrobial activity (Estrela et al., 2002). NaOCl is most widely used for water disinfection and can be used in disinfection of a clinical environment and industrial settings as food, pharmaceuticals, cosmetics and domestic products. NaOCl is an oxidizing biocide and exert its activity through the removal of electrons from sensitive functional groups and the targets can be the cell surface, cell wall or intracellular components giving these agents a very broad spectrum of activity (Estrela et al., 2002). However, NaOCl has some disadvantages as strong odour, decreased efficacy in the presence of organic material, unstable and prone to degradation resulting in formation of toxic by-products (Fukuzaki, 2006) A significant decrease in biofilm formation by S. Typhimurium and S. Enteritidis at all AgNPs concentrations was achieved and the anti-biofilms efficiency increased with the increase of concentration, this result agreed with that of Fabrega et al. (2009); Kaoud and Yosseif (2013) and Martinez-Gutierrez et al. (2013). In the current study, there were no significant differences between biofilm attenuation/prevention effect of various concentration including 10 ug/ml (sub MIC), 15 ug/ml (MIC), 20 ug/ml (MBC) and 25 ug/ml, indicating that sub MIC dose of silver does not exhibit cell killing or growth arresting activities but only interfere with biofilm forming ability. The attenuation in biofilm formation could be considered as a potential way to make the bacterial population more susceptible to antimicrobial agents so that they can be removed from the target site (Bjarnsholt et al., 2013). The great efficiency of AgNPs on the prevention of biofilm formation could be explained when the bacterial cells were treated with silver, they attached less to abiotic surfaces and lose their ability to form biofilm (Sharma et al., 2015). Moreover, the antibiofilm activity of AgNPs could be due to inhibition of EPS synthesis that reduces the biofilm formation (McLaughlin-Borlace et al. 1998). NaOCl was able to promote a significant reduction on the biofilm formation and its efficiency was concentration dependent. This result was in line with the results reported by Russell and McDonnell (2000). During treatment, NaOCl decomposes into sodium hydroxide and hypochlorite, which is a strong oxidizing agent (Tote et al., 2010) that act by oxidizing the cell membrane of bacterium resulting in cell lysis and death (Maris, 1995; Hawkins and Davies, 1999). Although, the chlorinated agents can penetrate into the bacterial biofilm, but these chemicals were not able to completely inactivate all bacteria because the microorganisms in biofilms carry protective mechanisms against the lethal effect of this type of biocidal agents (Stewart et al., 2001). Studying the effect of AgNPs on biofilms is important because bacteria are often present in biofilm communities. Compared to planktonic cells, biofilm cells have specific biological activities, metabolic pathways and stress responses (Stewart and Franklin, 2008). The EPS of biofilms act as a supporting structure for bacterial adherence and access to nutrients, as well as protects against antimicrobial agents (Høiby et al., 2010). The use of various antimicrobial agents as glutaraldehyde, formaldehyde and peroxygen at 1.0% under field conditions is unsatisfactory to eradicate Salmonella biofilms (Marin et al., 2009). The present study revealed that AgNPs were able to remove Salmonella spp. biofilm and increasing the contact time between AgNPs and biofilms increased the antibiofilm efficiency and this agreed with Rai et al. (2009). AgNPs are more destructive to biofilms due to better penetration into the EPS matrix (Habash et al., 2014; González et al., 2015). Mature biofilms can tolerate AgNPs by using EPS-mediated trapping, aggregation, and reduced diffusion (Peulen and Wilkinson, 2011; Sheng and Liu, 2011; Joshi et al., 2012). The increased resistance of mature biofilms to antimicrobial agents may be due to several factors. Firstly, bacterial cells in mature biofilms are in the stationary growth phase, therefore, less susceptible to antimicrobial agents (Anderl et al., 2003). Secondly, cells that die in the outer layers of mature biofilms could provide nutrients that enhance the growth of cells in deeper layers (Ito et al., 2009). Thirdly, the high thickness or high amount of EPS in mature biofilms may limit the transport of AgNPs through biofilms. Finally, mature biofilms produce not only more EPS but also other components as, curli which is a protein component used for bacterial adhesion to surfaces (Saldaña et al., 2009). Nevertheless, our results revealed that starch-stabilized AgNPs exhibited not only potent bactericidal activity, but also inhibit the biofilm formation as well as an efficient removal of the mature biofilm formed by S. Typhimurium and S. Enteritidis. NaOCl effectively eradicated the mature biofilms formed by S. Typhimurium and S. Enteritidis. Park et al. (2012) found that 100 mg/L NaOCl resulted in a ~1 log10 reduction in the number of cells from Salmonella biofilm recovered from steel cells after 5-50 min exposure. Ramesh et al. (2002) mentioned that 0.05% NaOCl was effective and completely eliminated Salmonella biofilm as well as produced 7.18 logarithmic reductions in Salmonella populations within 2 min., Moretro et al. (2009) found that NaOCl at concentrations of 1300 ppm completely eradicated Salmonella biofilms on polystyrene pegs after 1 min of exposure. Vestby et al. (2009) found a 2.4 log10 reduction of 48 h formed Salmonella enterica biofilm after treatment by 500 mg/l NaOCl for 5 min. Additionally, Wong et al. (2010) found that 1 min. exposure S. Typhimurium biofilms (3 days old) to NaOCl (1.31 g/l) reduced the bacteria to undetectable levels. Increasing the disinfectant concentration and contact time increased its effectiveness against biofilms (Russell and McDonnell, 2000). Biofilms protects bacteria from detergents and sanitizers, and the bacterial cells are able to develop specific attributes like low multiplication rates or additional defense mechanisms; due to the ability of the extracellular polymeric matrix to neutralize antimicrobial agents, because it consists of organic matter (Rossi and Porto, 2009). Furthermore, mature biofilm provides a more protective factor for bacteria against bactericidal agents. The present study showed that mature biofilms formed by S. Typhimurium and S. Enteritidis were significantly reduced after treatment with NaOCL. Conclusion AgNPs and NaOCl were able to promote significant reduction and control the mature biofilms of S. Typhimurium and S. Enteritidis isolated from poultry farms. However, the use of AgNPs has the ability to overcome NaOCl disadvantages as odour, instability and decreased efficacy in the presence of organic material. AgNPs is superior to many other antimicrobial agents as they have not been shown to cause bacterial resistance. From these, it can be suggested that AgNPs can be embedded into the matrices or material used for the fabrication of medical devices and poultry equipment to avoid adherence and formation of microbial biofilms. The study also recommends AgNPs compounds to be used as sanitary wipes in food industry and food-related activities. Further research and development are necessary to translate that technology into preventive and therapeutic strategies. NaOCL was also effective in prevention and controlling of biofilms formed by S. Typhimurium, S. Enteritidis but at higher concentration compared by AgNPs and the results magnified the antibiofilm role of AgNPs compared with NaOCL as 25 mg/l AgNPs was more effective to prevent biofilms than 2200 mg/l NaOCL. So, we can consider AgNPs as a new category of disinfectants that can affect both planktonic and sessile form of bacteria in an excellent manner. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Conflict of Interests |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Authors declare no conflicts of interest to disclose. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
References |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abd El-Ghany, W.A., El-Shafii, S.S., Hatem, M.E., 2012. A survey on Salmonella species isolated from chicken flocks in Egypt. Asian Journal of Animal and Veterinary Advances 7(6), 489-501.þ Abidi, S.H., Ahmed, K., Sherwani, S.K., Kazmi, S. U., 2014. Reduction and removal of Pseudomonas aeruginosa biofilm by natural agents. Int. J. Chem. Pharm. Sci. 5, 28-34.þ Anderl, J.N., Zahller, J., Roe, F., Stewart, P.S., 2003. Role of nutrient limitation and stationary-phase existence in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrobial Agents and Chemotherapy 47(4), 1251-1256.þ Anon, 2007. Microbiology of food and animal feeding stuffs. Horizontal method for the detection of Salmonella spp. Annex D: detection of Salmonella spp. in animal faeces and in environmental samples from the primary production stage. EN ISO 6579: 2002/Amd1. International Organization for Standardization, Geneva. Anonymous, 2004. Morbidité et mortalité dues aux maladies infectieuses d'origine alimentaire en France. In: Rapport Institut de Veille Sanitaire - Agence Française de Sécurité Sanitaire des Aliments. Antunes, P., Réu, C., Sousa, J. C., Peixe, L., Pestana, N., 2003. Incidence of Salmonella from poultry products and their susceptibility to antimicrobial agents. International Journal of Food Microbiology 82(2), 97-103.þ Bjarnsholt, T., Ciofu, O., Molin, S., Givskov, M., Hoiby, N., 2013. Applying Insights from Biofilm Biology to Drug Development—Can a New Approach Be Developed?. Nature Reviews Drug Discovery 12, 791-808. Bogino, P.C., Oliva, M. D.L.M., Sorroche, F.G., Giordano, W., 2013. The role of bacterial biofilms and surface components in plant-bacterial associations. International Journal of Molecular Sciences 14(8), 15838-15859.þ Capita, R., Buzón-Durán, L., Riesco-Peláez, F., Alonso-Calleja, C., 2017. Effect of Sub-Lethal Concentrations of Biocides on the Structural Parameters and Viability of the Biofilms Formed by Salmonella Typhimurium. Foodborne Pathogens and Disease 14(6), 350-356. Carrique-Mas, J.J., Breslin, M., Snow, L., McLaren, I., Sayers, A.R., Davies, R.H., 2009. Persistence and clearance of different Salmonella serovars in buildings housing laying hens. Epidemiology and Infection 137(6), 837-846.þ Cevallos-Cevallos, J.M., Gu, G., Danyluk, M.D., van Bruggen, A.H., 2012. Adhesion and splash dispersal of Salmonella enterica Typhimurium on tomato leaflets: effects of rdar morphotype and trichome density. International Journal of Food Microbiology 160(1), 58-64. Coffey, B.M., Anderson, G.G., 2014. Biofilm formation in the 96-well microtiter plate. Methods Mol Biol. 1149, 631-41 Corcoran, M., Morris, D., De Lappe, N., O’Connor, J., Lalor, P., Dockery, P., Cormican, M. (2014): Commonly used disinfectants fail to eradicate Salmonella enterica biofilms from food contact surface materials. Appl. Environ. Microbiol. 80, 1507–1514. Cos, P.; Toté, K.; Horemans, T., Maes, L., 2010. Biofilms: an extra hurdle for effective antimicrobial therapy. Current Pharmaceutical Design, 16, 2279-2295 Donskey, C.J., 2013. Does improving surface cleaning and disinfection reduce health care-associated infections? American Journal of Infection Control 41(5), S12-S19. Espigares E., Bueno A., Espigares M., Ga ́lvez R., 2006. Isolation of Salmonella serotypes in wastewater and effluent: Effect of treatment and potential risk. Int. J. Hyg. Environ.-Health 209, 103–107 Estrela C.L., Estrela C.R., Barbin E.L., Spanó J.C., Marchesan M.A., Pécora J.D., 2002. Mechanism of action of sodium hypochlorite. Braz. Dent. J. 13(2), 113-117. Fabrega, J., Renshaw, J.C., Lead, J.R., 2009. Interactions of silver nanoparticles with Pseudomonas putida biofilms. Environmental Science and Technology 43(23), 9004-9009. Fabrega, J., Luoma S.N., Tyler, C.R., Galloway T.S., Lead J.R., 2011. Silver nanoparticles: Behaviour and effects in the aquatic environment. Environment International 37, 517–531 Fukuzaki, S., 2006. Mechanisms of Action of Sodium Hypochlorite in Cleaning and Disinfection Processes. Biocontrol Science 11, 147-157. Ganjali Dashti, M., Abdeshahian, P., Sudesh, K., Phua, K.K., 2016. Optimization of Salmonella Typhi biofilm assay on polypropylene microtiter plates using response surface methodology. Biofouling, 32(4), 477-487. Gnanadhas D.P., Thomas M.B., Thomas R., Raichur A.M, Chakravortty D., 2013. Interaction of silver nanoparticles with serum proteins affects their antimicrobial activity in vivo. Antimicrobial Agents Chemotherapy 57, 4945-4955. González, A.G., Mombo, S., Leflaive, J., Lamy, A., Pokrovsky, O.S., Rols, J.L., 2015. Silver nanoparticles impact phototrophic biofilm communities to a considerably higher degree than ionic silver. Environmental Science and Pollution Research 22(11), 8412-8424.þ Habash, M.B., Park, A.J., Vis, E.C., Harris, R.J., Khursigara, C.M., 2014. Synergy of silver nanoparticles and aztreonam against Pseudomonas aeruginosa PAO1 biofilms. Antimicrobial agents and Chemotherapy 58(10), 5818-5830. Hawkins, C.L., Davies, M.J., 1999. Hypochlorite-induced oxidation of proteins in plasma: formation of chloramines and nitrogen-centred radicals and their role in protein fragmentation. Biochemical Journal 340 (2), 539-548.þ Hilbert F., Smulders F.J.M., Chopra-Dewasthaly, R., Paulsen, P., 2012. Salmonella in the wildlife-human interface. Salmonella in Foods: Evolution, Strategies and Challengeies 45, 603-608. Hoelzer, K., Switt, A.I.M., Wiedmann, M., 2011. Animal contact as a source of human non-typhoidal salmonellosis. Veterinary Research 42, 34.þ Høiby, N., Bjarnsholt, T., Givskov, M., Molin, S., Ciofu, O., 2010. Antibiotic resistance of bacterial biofilms. International Journal of Antimicrobial Agents 35(4), 322-332.þ Holt, K.B., Bard, A.J., 2005. Interaction of silver (I) ions with the respiratory chain of Escherichia coli: An electrochemical and scanning electrochemical microscopy study of the anti- microbial mechanism of micromolar Ag+. Biochemistry 44, 13214–13223. Irayyif, S.M., Mohseni, A., Araghi, S., Degree, I.A., 2015. Silver nanoparticles and their effect on the biofilm formation in food borne Salmonella species. International Journal of Recent Scientific Research 6, 4343-4346. Ito, A., Taniuchi, A., May, T., Kawata, K., Okabe, S., 2009. Increased antibiotic resistance of Escherichia coli in mature biofilms. Applied and Environmental Microbiology 75(12), 4093-4100.þ Janssens, J.C., Steenackers, H., Robijns, S., Gellens, E., Levin, J., Zhao, H., Vanderleyden, J., 2008. Brominated furanones inhibit biofilm formation by Salmonella enterica Serovar Typhimurium. Applied and Environmental Microbiology 74(21), 6639-6648. Jassim, S.A.A., Limoges, R.G., 2017. Bacteriophage biocontrol in poultry. In: Bacteriophages: Practical Applications for Nature's Biocontrol. Springer, Cham. Joshi, N., Ngwenya, B.T., French, C.E., 2012. Enhanced resistance to nanoparticle toxicity is conferred by overproduction of extracellular polymeric substances. Journal of Hazardous Materials 241, 363-370.þ Kanematsu, H., Ikigai, H., Yoshitake, M., 2009. Evaluation of various metallic coatings on steel to mitigate biofilm formation. International Journal of Molecular Sciences 10(2), 559-571 Kaoud, H.A., Yosseif, S., 2013. Efficacy of silver nanoparticles and activated electro- chemical water as poultry disinfectants against Salmonella Enteritidis 1, 8–13. Li, Y., Leung, P., Yao, L., Song, Q. W., Newton, E., 2006. Antimicrobial effect of surgical masks coated with nanoparticles. Journal of Hospital Infection 62(1), 58-63. Loo Y. Y., Rukayadi Y., Nor-Khaizura M.-R., Kuan C. H., Chieng B. W., Nishibuchi M., Radu S., 2018. In Vitro Antimicrobial Activity of Green Synthesized Silver Nanoparticles Against Selected Gram-negative Foodborne Pathogens. Frontiers in Microbiology 9, 1-7 Mallinson, E.T., de Rezende C.E., Tablante NL., Carr L.E., Joseph S.W., 2000. A management technique to identify prime of Salmonella contamination on broiler and layer farms. J. Appl. Poult. Res. 9, 364-370. Marin, C., Hernandiz, A., Lainez, M., 2009. Biofilm development capacity of Salmonella strains isolated in poultry risk factors and their resistance against disinfectants. Poultry Science 88(2), 424-431.þ Maris P., 1995. Mode of Action of Disinfectants. Scientific and Technical Review of the Office International des Epizooties (Paris) 14, 47-55 Martinez-Gutierrez, F., Boegli, L., Agostinho, A., Sánchez, E.M., Bach, H., Ruiz, F., James, G., 2013. Anti-biofilm activity of silver nanoparticles against different microorganisms. Biofouling 29(6), 651-660.þ McLaughlin-Borlace, L., Stapleton, F., Matheson, M., Dart, J.K.G., 1998. Bacterial biofilm on contact lenses and lens storage cases in wearers with microbial keratitis. Journal of Applied Microbiology 84(5), 827-838. Meerburg B.G., Kijlstra A., 2007. Role of rodents in transmission of Salmonella and Campylobacter. Journal Science Food Agriculture 87, 2774- 2781. Moretro, T., Vestby, L.K., Nesse, L.L., Storheim, S., Kotlarz, K., Langsrud, S., 2009. Evaluation of efficacy of disinfectants against Salmonella from the feed industry. Journal of Applied Microbiology 106(3), 1005-1012.þ Morones, J.R., Elechiguerra, J.L., Camacho, A., Holt, K., Kouri, J.B., Ramírez, J.T., Yacaman, M.J., 2005. The bactericidal effect of silver nanoparticles. Nanotechnology 16(10), 2346-2353.þ Murray, C.J., 2000. Environmental Aspects of Salmonella. Salmonella in Domestic Animals. Wray, C. and Wray, A. editors. CABI Publishing, New York, NY. Nilsson, R.E., Ross, T., Bowman, J.P., 2011. Variability in biofilm production by Listeria monocytogenes correlated to strain origin and growth conditions. International Journal of Food Microbiology 150, 14–24. Ogden, I.D., MacRae, M., Johnston, M., Strachan, N.J.C., Cody, A.J., Dingle, K.E., Newell, D.G., 2007. Use of multilocus sequence typing to investigate the association between the presence of Campylobacter spp. in broiler drinking water and Campylobacter colonization in broilers. Applied and Environmental Microbiology 73(16), 5125-5129.þ Oliveira, F.A., Pasqualotto, A.P., da Silva, W.P., Tondo, E.C., 2012. Characterization of Salmonella Enteritidis isolated from human samples. Food Research International 45(2), 1000-1003.þ Pal, S., Tak, Y.K., Song, J.M., 2007. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the gram-negative bacterium Escherichia coli. Applied and Environmental Microbiology 73(6), 1712-1720.þ Panáček, A., Kvítek, L., Prucek, R., Kolář, M., Večeřová, R., Pizúrová, N., Sharma, V. K., Nevěčná, T.J., Zbořil, R., 2006. Silver Colloid Nanoparticles: Synthesis, Characterization, and Their Antibacterial Activity. The Journal of Physical Chemistry B. 110, 16248-16253. Park, S.H., Cheon, H.L., Park, K.H., Chung, M.S., Choi, S.H., Ryu, S., Kang, D. H., 2012. Inactivation of biofilm cells of foodborne pathogen by aerosolized sanitizers. International Journal of Food Microbiology 154(3), 130-134.þ Peulen, T.O., Wilkinson, K.J., 2011. Diffusion of nanoparticles in a biofilm. Environmental Science and Technology 45(8), 3367-3373.þ Pui, C.F., Wong, W.C., Chai, L. C., Tunung, R., Jeyaletchumi, P., Hidayah, N., Son, R., 2011. Salmonella: A foodborne pathogen. International Food Research Journal 18(2). Rai, M., Yadav, A., Gade, A., 2009. Silver nanoparticles as a new generation of antimicrobials. Biotechnology Advances 27(1), 76-83 Ramesh, N., Joseph, S.W., Carr, L.E., Douglass, L.W., Wheaton, F.W., 2002. Evaluation of chemical disinfectants for the elimination of Salmonella biofilms from poultry transport containers. Poultry Science 81, 904-910. Rodrigues, D., Cerca, N., Teixeira, P., Oliveira, R., Ceri, H., Azeredo, J., 2011. Listeria monocytogenes and Salmonella enterica Enteritidis biofilms susceptibility to different disinfectants and stress-response and virulence gene expression of surviving cells. Microbial Drug Resistance 17(2), 181-189 Rodriguez, A., Pangloli, P., H.A. Richards, J.R. Mount, Draughon, F.A. 2006. Prevalence of Salmonella in diverse environmental farm samples. J. Food Prot. 69, 2576– 2580. Rossi A.C.R., Porto E., 2009. A importância da elaboração de procedimen¬tos dehigienização considerando a presença de biofilmes. Sociedade Brasileira de Controle de Contaminação. Março/Abril 2009, pp.40-41. Russell, A.D., McDonnell, G., 2000. Concentration: a major factor in studying biocidal action. Journal of Hospital Infection 44(1), 1-3.þ Saldaña, Z. Xicohtencatl‐Cortes, J., Avelino, F., Phillips, A.D., Kaper, J.B., Puente, J.L., Girón, J.A., 2009. Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli. Environmental Microbiology 11(4), 992-1006.þ Samberg M.E., Orndorff, P.E., Monteiro-Riviere, N.A., 2011. Antibacterial efficacy of silver nanoparticles of different sizes, surface conditions and synthesis methods. Nanotoxicology 5(2), 244–253 Sharma, B.K., Saha, A., Rahaman, L., Bhattacharjee, S., Tribedi, P., 2015. Silver inhibits the biofilm formation of Pseudomonas aeruginosa. Advances in Microbiology 5(10), 677-685.þ Sheffield, C.L., Crippen, T.L., 2012. Invasion and survival of Salmonella in the environment: The role of biofilms. In Salmonella-A Diversified Superbug. InTech. Shemesh, M., Tam, A., Steinberg, D., 2007. Differential gene expression profiling of Streptococcus mutans cultured under biofilm and planktonic conditions. Microbiology, 153(5), 1307-1317.þ Sheng, Z., Liu, Y., 2011. Effects of silver nanoparticles on wastewater biofilms. Water research, 45(18), 6039-6050.þ Shrivastava, S., Bera, T., Roy, A., Singh, G., Ramachandrarao, P., Dash, D., 2007. Characterization of enhanced antibacterial effects of novel silver nanoparticles. Nanotechnology 18(22), 225103-225112 Soumet, C., Ermel, G., Rose, N., Rose, V., Drouin, P., Salvat, G., Colin, P., 1999. Evaluation of a multiplex PCR assay for simultaneous identification of Salmonella spp., Salmonella Enteritidis and Salmonella Typhimurium from environmental swabs of poultry houses. Letters in Applied Microbiology 28(2), 113-117.þ Stewart, P.S., Rayner, J., Roe F., Rees W.M., 2001. Biofilm penetration and disinfection efficacy of alkaline hypochlorite and chlorosulfamates. J. Appl. Microbiol. 91(3), 525-532 Stewart, P.S., Franklin, M.J., 2008. Physiological heterogeneity in biofilms. Nature Reviews Microbiology 6(3), 199–210. Tote, K., Horemans, T., Berghe, D.V., Maes, L., Cos, P., 2010. Inhibitory effect of biocides on the viable masses and matrices of Staphylococcus aureus and Pseudomonas aeruginosa biofilms. Applied and Environmental Microbiology 76(10), 3135-3142. Vestby, L.K., Moretro, T., Ballance, S., Langsrud, S., Nesse, L.L., 2009. Survival potential of wild type cellulose deficient Salmonella from the feed industry. BMC Veterinary Research 5, 43. Vigneshwaran, N., Nachane, R.P., Balasubramanya, R.H., Varadarajan, P.V., 2006. A novel one-pot ‘green’synthesis of stable silver nanoparticles using soluble starch. Carbohydrate Research 341(12), 2012-2018.þ Wales, A., Breslin, M., Davies, R., 2006. Semiquantitative assessment of the distribution of Salmonella in the environment of caged layer flocks. Journal of Applied Microbiology, 101(2), 309-318.þ Wilks, S.A., Michels, H.T., Keevil, C.W., 2006. Survival of Listeria monocytogenes Scott A on metal surfaces: implications for cross-contamination. International Journal of Food Microbiology 111(2), 93-98.þ Wong, H.S., Townsend, K.M., Fenwick, S.G., Trengove, R.D., O’handley, R.M., 2010. Comparative susceptibility of planktonic and 3‐day‐old Salmonella Typhimurium biofilms to disinfectants. Journal of Applied Microbiology 108(6), 2222-2228.þ Yezli, S., Otter, J.A., 2012. Does the discovery of biofilms on dry hospital environmental surfaces change the way we think about hospital disinfection? Journal of Hospital Infection 81(4), 293-294. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||